Leishmania are dimorphic parasites that present as two principal morphological stages: the intracellular amastigote, in the cells of the mammalian host mononuclear phagocyte system, and the flagellated promastigote, in the intestinal tract of the insect vector. Since the first Leishmania species was described (Laveran and Mesnil, 1903), the number of species has increased steadily, and currently stands around 30. As the different species are morphologically indistinguishable, other characteristics have been used for their taxonomy. Although DNA-based methods are increasingly being used, isoenzyme electrophoresis remains the gold standard technique for identification and is the basis for the current classification (Table 1).

Vector
Sandflies are psychodid Diptera of the subfamily Phlebotominae. Their life cycle includes two different biological stages: the free-flying adult and the developmental stages, which include egg, four larval instars, and pupa, and occur in damp soil rich in organic material. The adults are small flying insects about 2–4 mm in length, with a yellowish hairy body. During day, they rest in dark, sheltered places. They are active at dusk and during the night. Both sexes feed on plant juices, but females also need a blood meal before they are able to lay eggs. It is during this blood meal that Leishmania parasites are transmitted between the mammalian hosts. Among the 800 known species of sand flies, about 70, belonging to the genera Phlebotomus in the Old World and Lutzomyia in the New World, are proven or suspected vectors of Leishmania, and a certain level of specificity exists between Leishmania and sand fly species.Reservoir
Various species of seven different orders of mammals are the reservoir hosts responsible for long-term maintenance of Leishmania in nature. Depending on the focus, the reservoir host can be either a wild or a domestic mammal, or even in particular cases human beings. In visceral leishmaniasis, these different types of reservoir host represent different steps on the hypothetical path toward ‘anthropization’ of a ‘wild’ zoonosis (Garnham, 1965). Rodents, hyraxes, marsupials, and edentates are common reservoirs of wild zoonotic CL. Dogs are currently considered as true reservoirs of L. infantum and L. peruviana, two species that have peri-domestic or even domestic transmission. Humans are the commonly recognized reservoir of L. donovani VL and L. tropica CL.Life Cycle And Transmission
In nature, Leishmania parasites are alternately hosted by the insect (flagellated promastigote) and by mammals (intracellular amastigote stage). Leishmaniasis is normally transmitted to humans by the inoculation of metacyclic promastigotes through the sand fly bite. Other routes remain exceptional. Exchange of syringes is thought to explain the high prevalence of L. infantum/human immunodeficiency virus (HIV) co-infection in intravenous drug users in southern Europe (Alvar and Jimenez, 1994).Geographical Distribution
Leishmaniasis occurs in more than 88 countries, ranging over the intertropical zones of America and Africa, and extending into temperate regions of South America, southern Europe, and Asia. The limits of the disease are latitudes 45 north and 32 south. The geographical distribution is governed by those of the mammal or sand fly host species, their ecology, and their own distribution area. The leishmaniases are diseases with natural focality. They include several ‘noso-epidemiological units,’ which can be defined as the conjunction of a particular Leishmania species circulating in specific natural hosts, evolving in a natural focus with specific ecological patterns, and having a particular clinical expression (see Table 2).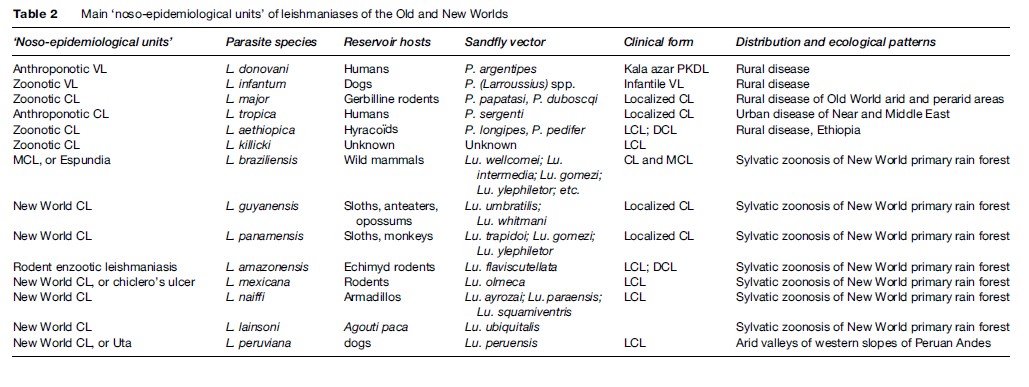
Visceral Leishmaniasis
The two viscerotropic species of Leishmania have distinct life cycles and geographic distribution. The anthroponotic species L. donovani is restricted to India and East Africa. The disease is known as ‘kala-azar’ and can be complicated by a chronic CL form called ‘post-kala-azar dermal leishmaniasis.’ The zoonotic species L. infantum extends from China to Brazil, and is responsible for infantile VL. The main historical foci of endemic VL are located in China, India, Central Asia, East Africa, the Mediterranean basin, and Brazil.Old World Cutaneous Leishmaniasis
The large majority of Old World CL cases are due to the species L. major or L. tropica and occur mainly in countries of the Near and Middle East: Afghanistan, Iran, Saudi Arabia, and Syria. L. major is responsible for zoonotic CL, of which reservoir hosts are gerbilline rodents, and vectors are sand flies of the subgenus Phlebotomus, principally Phlebotomus papatasi and P. duboscqi. This species has a wide distribution, including west, north, and east Africa, the Near and Middle East, and Central Asia. L. tropica is an anthroponotic species with humansacting as reservoir hosts and the vector being P. (Paraphlebotomus) sergenti. It occurs in various cities of the Near and Middle East, but extends also to Morocco, where the dog is suspected to be a reservoir host in some foci. Other species have restricted distributions: L. aethiopica in Ethiopia and Kenya, and L. killicki in Tunisia.New World Tegumentary Leishmaniasis
In the New World, L. braziliensis is the species responsible for MCL. It has a wide distribution, extending from southern Mexico to northern Argentina. L. amazonensis has a wide South American distribution, but human cases of this rodent enzootic species are unusual. Other species have more restricted distributions: L. guyanensis (north of the Amazon basin), L. panamensis (Colombia and Central America), L. mexicana (Mexico and Central America), and L. peruviana, which is restricted to Andean valleys of Peru. With the exception of this last species, all American dermotropic species are responsible for sylvatic zoonoses occurring within the rain forest.Disease Burden
An estimated 350 million people in more than 88 countries on four continents are ‘at risk’ of leishmaniasis. The annual incidence of new cases, including all clinical forms, is estimated between 1.5 and 2 million (Desjeux, 1999). Differences in morbidity, or even in mortality, depend on the form of the disease. The clinical expression of leishmaniasis depends not only on the genetically determined tropism of the different Leishmania species (viscero-, dermo-, or mucotropisms) but also on the immunological status of the infected patient.
